| HOME |

|
|
|
Cephalia: Juvenile/Adult Phase
Change in Cacti Apical cephalia Like most plants, cactus seedlings are unable to flower immediately after germination. They must grow as juveniles for a period of several months to many years before they can finally be induced to flower by environmental or internal conditions; once they become inducible, they are adults, whether actually flowering or not. Also like most plants in general, most cacti undergo little or no noticeable change in growth pattern once they undergo their phase change from juvenile to adult condition: an old, adult plant capable of flowering looks just like a young, juvenile plant.
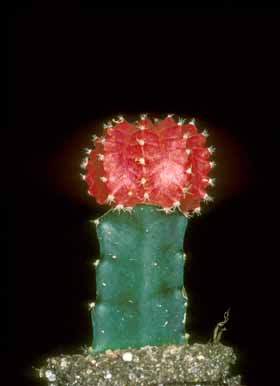
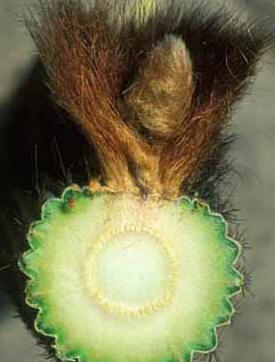
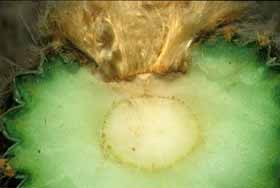
However, in several genera, the phase change is accompanied by a dramatic change in growth pattern and body form, one that is much more striking than occurs in any other group of plants. The adult body form differs so greatly from the juvenile that many people – many botanists – assume that the plant is actually two separate genera grafted together. Excellent examples occur in Melocactus. All species of Melocactus grow initially as globose cacti with spherical, green bodies with about 8 to 27 ribs, and with axillary buds whose strong, curved spines are spaced widely enough that they protect the shoot but do not shade it significantly. The plants grow like this for years without ever flowering. Once old enough (we do not know if the stimulus is age, size or some other factor), the plants switch to being adults and alter their growth. All Melocactus plants are unbranched, so the adult phase body is produced by the same shoot apical meristem as the juvenile phase body. The adult phase body is much narrower than that of the juvenile because both pith and cortex are narrower. The surface of the adult stem is smooth, lacking ribs and tubercles, and phyllotaxy is altered to be very high such that axillary buds are produced very close together. Spine morphology too changes, with spines being shorter and straighter and so abundant that – combined with the close packing of the axillary buds – the adult surface looks like a brush with tightly packed bristles. Although there is virtually no space at all between spines, somehow the axillary buds also produce prodigious amounts of trichomes: the combination of trichomes and spines completely protects the adult surface from animals, sunlight, fungi, bacteria, even from plant anatomists. With no light whatsoever penetrating the mass of spines, the adult phase cortex completely lacks both chlorophyll and a palisade cortex, although it does have cortical bundles. With no chlorophyll there is also no need for stomata, and with no need for stomata there is no need for an epidermis: epidermis cells convert to acting as cork cambium cells immediately below the zone where leaf primordia and axillary buds are formed. The adult shoot never has a mature epidermis, only a protoderm at the shoot apex and bark on all other surfaces. This adult phase body is called a cephalium, and because it is located at the upper end of the shoot, it is an apical cephalium.
As mentioned above, no other family of plants has members that undergo such a striking phase change. Of course, many insects do, with their metamorphosis from a caterpillar to a butterfly, but that involves a rearrangement of existing tissues and the animal has its two bodies in sequence. In Melocactus and other cacti with apical cephalia, the tissues of the juvenile body are not rearranged, it is just that all new adult phase growth is added on to the top of the juvenile body. Within the body, the vascular cambium undergoes a phase change as well, with the result that all wood in the cephalium is WBT wood, and all new wood produced in the juvenile body after the phase change is WBT wood as well (the vascular cambium in the juvenile body had produced fibrous wood while the plant consisted of only the juvenile phase). The adult phase body provides wonderful protection for developing flowers and fruits. Apical meristems of axillary buds of the adult phase become active and begin producing flower appendage primordia; while doing this, they are located beneath one or more centimeters of impenetrable spines and trichomes. Once the flower is mature and ready for anthesis (ready to open), all cells just above the ovary absorb water and swell, the flower expands enough to poke the petal tips out beyond the spines tips, then the petal tips curve sideways, creating a nice smooth covering over and between the spines. As a pollinator visits the flower, it can insert its proboscis through a silky tube down to the nectaries at the base of the stamens with no risk at all of being impaled on any of the spines. The emergence of the flowers is dramatic: it happens overnight and one can check a plant in the evening with no hint of flowers at all, then the next morning the bright red flowers are completely open. Flowers last only a single day, then the petals wither, blocking off all access to the ovary. The ovules mature into seeds and the ovary matures into a fruit, completely hidden and protected among the bases of all those spines. Once mature, again there is an absorption of water and a rapid swelling but this time by cells at the base of the ovary such that the ovary (now a seed-filled fruit) is pushed up to the top of the spines where it is visible to fruit-eating animals that might disperse the seeds. The protection offered by the adult body is remarkable, but there is another benefit to this juvenile/adult transition that may be equally important. The morphology of the adult phase makes it less expensive to produce a flower. Like many plants, most cacti produce only a single flower from each axillary bud, so if a plant needs to make 20 flowers, it must make 20 axillary buds beforehand. For ordinary plants with slender twigs, this probably is not a significant cost: the slender internodes are not expensive and if the plant does not need 20 more leaves, it can allow the axillant leaf primordia to develop as minute, cheap scales. But for a cactus with a broad stem many centimeters in diameter, making extra internodes is costly: to make 20 flowers, it will have to make 20 internodes worth of voluminous cortex, 20 internodes worth of epidermis through which it might lose more water and so on. This is acceptable if the plant does need more water-storage tissue and more photosynthetic surface area, but the evolution of highly modified stem-succulence has linked flowering/photosynthesis/water conservation/building costs into a tight relationship that might not allow any of those processes to become optimized evolutionarily. But evolution of the cephalium broke that tight relationship: the adult phase has narrower shoots and more closely packed nodes and axillary buds, which greatly reduces the cost of building each new axillary bud and thus each new flower. But the positive benefits of having an apical cephalium are accompanied by negative consequences as well. Once each plant switches from the juvenile to the adult phase, it becomes incapable of producing any new chlorophyllous, photosynthetic tissue. After reaching adulthood, not only is the total amount of photosynthetic tissue set, but it begins to age as well. It could be that the existing chlorenchyma cells can somehow recycle their chloroplasts, producing new ones while breaking down old ones, but it is not known if that actually happens. We do know for certain that the cells themselves are not replaced: those in the juvenile portions of very old plants look like old cells. It would seem that with each passing year, the photosynthetic capacity of the plant would decrease, but in nature there are plants with extremely tall cephalia which appear to have lived for many, many years in the adult phase. The juvenile/adult transition can be reversed. If an old plant with a cephalium is decapitated by cutting off either the top of the cephalium or the entire cephalium, a dormant axillary bud will become activated and grow out, but always with the juvenile morphology. After growing for a brief period and while rather small, the bud will begin to make a cephalium. If just the top of the cephalium had been removed, the activated bud is one of the cephalium buds, so the plant consists of (from ground upward), original juvenile, base of cephalium, new juvenile bud, new cephalium. If the original decapitation had removed all the cephalium, then of course it is an axillary bud of the juvenile that becomes active, and it too initially has juvenile morphology but quickly produces a cephalium. Once again we do not know what controls the size the bud needs to achieve before it can start making its own cephalium. Perhaps the bud must grow until its volume equals that of the part that had been removed by decapitation. A species that appears ideal for this research is Melocactus intortus: these are small plants which, under ideal conditions in a greenhouse with lots of fertilizer, initiate their cephalia while only 3 or 4 years old. Apical cephalia occur also on Discocactus and Backebergia. Discocacti are small, globose cacti like melocacti, but plants of Backebergia (which have been studied by Teresa Terrazas and Monserrat Vasquez Sanchez) are giant, highly branched columnar cacti. A shoot must become many meters tall before it undergoes its phase change and initiates its apical cephalium, and the phase change affects only that particular branch: other branches on the same plant continue growing as juveniles. After a cephalium has grown for many years, the plant abscises it and throws it away, and the ground below a large plant is littered with decomposing cephalia. An axillary bud just below the abscission zone produces a new juvenile branch, and this grows for many years as a photosynthetic juvenile before switching to adulthood. Backebergia has found a way to continue to make photosynthetic tissue even after it has started to reproduce.
Mauseth, J. D. 1989. Comparative structure-function studies within a single strongly dimorphic species, Melocactus intortus (Cactaceae). Bradleya 7: 1 – 12. Mauseth, J. D., T. Terrazas, M. Vasquez-Sanchez, S. Arias. 2005. Field observations on Backebergia militaris and other cacti of the Balsas Basin, Mexico. The Cactus and Succulent Journal (USA) 77: 2 - 13.
Lateral cephalia There is another way to continue making photosynthetic tissue even after a cactus has started making its cephalium: make a lateral cephalium instead of an apical cephalium. This occurs in Espostoa, Pseudoespostoa, Thrixanthocereus, Vatricania (now all considered to be Espostoa), Buiningia, Facheiroa, Micranthocereus and others. These plants are all columnar (Buiningia aurea is such a short column it appears to be globose – it may grow quickly enough to be a good species for research) and grow as ribbed green juvenile columns with an ordinary sparse spine cover. Once old enough, they begin making a cephalium, but only along one side. A plant of Espostoa might have 18 to 25 ribs, but only several of them undergo the phase change: as the shoot apical meristem adds new cells to the upper ends of these special ribs, they develop only into a narrow cortex without chlorophyll but with long, abundant spines with copious trichomes, bark rather than epidermis, and the ribs develop so little they remain low and indistinct when mature. The axillary buds on these low ribs will be able to form flowers. Meanwhile, the new cells being added to the upper ends of all the rest of the ribs being produced by the very same shoot apical meristem continue to develop into tissues with juvenile characters. Of the numerous ribs produced by a single meristem, most develop with juvenile morphology, a few develop simultaneously with adult morphology. Recall when you went through puberty: imagine that only one strip of your body had changed while all the rest of you continued to grow with child-like features. That is what these plants with lateral cephalia do. They are able to produce photosynthetic juvenile tissues throughout their entire lives, even after part of their bodies are adult and flowering; they avoid the problem Melocactus faces.
Mauseth, J. D. 1999. Comparative anatomy of Espostoa, Pseudoespostoa, Thrixanthocereus, and Vatricania (Cactaceae). Bradleya 17: 33 – 43. Kiesling, R., and J D. Mauseth. 2000. History and taxonomy of Neoraimondia herzogiana (Cactaceae). Haseltonia 7: 47 – 52.
Cephalia in Neoraimondia There is still another solution to making it less expensive to flower without giving up the ability to make photosynthetic tissue. Plants of Neoraimondia (and the closely related Neocardenasia herzogiana) branch from the base and each branch then grows upward as a massive green cactus stem. The axillary buds of this green stem are unusual in being able to produce several flowers each year, as well as flowering year after year. Each time the axillary bud flowers, it first produces a tiny bit of stem with a few axillary buds of its own. These axillary buds themselves can flower, but they too first produce a bit of stem and more axillary buds. After a few years, all the bits of stem add up to a real branch. But this branch does not look at all like the original green photosynthetic branch that produced the original axillary bud. Instead, it is very narrow, has no ribs, no epidermis (just bark), no photosynthetic tissues and is covered so tightly with short spines and trichome that it resembles the inflorescence of a cattail (Typha). It is in effect a cephalium: the axillary buds of Neoraimondia grow out not as ordinary branches but as cephalia. The original green shoot can continue to grow and make more axillary buds, each of which will eventually grow out as a cephalium. A strange thing is that the green photosynthetic branches are determinate: they become very long and massive but eventually stop growing at a characteristic height. Then a bud at the base of the plant grows out as another green photosynthetic shoot, and its axillary buds too will eventually grow out as cephalia. Like Backebergia and Espostoa, plants of Neoraimondia can produce flowers cheaply while also producing new photosynthetic tissues.
The biology of Neoraimondia is very unusual and has many facets in need of study, but I do not know of any large plants in cultivation in the United States. These plants need a lot of room and must be cultivated with no risk of frost. Mauseth, J. D., and R. Kiesling. 1997. Comparative anatomy of Neoraimondia roseiflora and Neocardenasia herzogiana (Cactaceae). Haseltonia 5: 37 – 50. Kiesling, R., and J D. Mauseth. 2000. History and taxonomy of Neoraimondia herzogiana (Cactaceae). Haseltonia 7: 47 – 52. There is much more information about cephalium-bearing plants and the areas where they grow in A Cactus Odyssey. [end Cephalia page]
|